The electron transport chain (ETC) is a highly specialised and biochemical family of proteins (mitochondrial complexes I, II, III and IV) which work in concert with mitochondrial complex V (the ATP synthase), and accessory molecules (ubiquinone and cytochrome c) and are responsible for one of the main functions of mitochondria, that of the conversion of adenosine-5′-diphosphate (ADP) into adenosine- 5′-trisphosphate (ATP), which is molecule the common chemical energy currency of the cell. It is situated in the inner membrane of the eukaryotic mitochondrion. The pathway itself is composed of four separate enzymatic complexes that act in sequence to allow the function of a fifth complex; ATP synthase. Through the process of oxidative phosphorylation, the enzymes and accessory molecules in this pathway, in co-operation with ATP synthase the proverbial “heart” of the mitochondrion, for it is this component that is singularly responsible for the production of adenosine triphosphate (ATP); the chemical energy source that drives a truly massive array of cellular processes. Each ETC component (as well as ATP synthase) has its own specific structure and function/s and as such our series of posts entitled ‘The ETC Disassembled’ will focus on each individual component of the ETC, complex I, complex II, complex III, complex IV and complex V, as well the accessory electron carriers, ubiquinone (CoQ10) and cytochrome c, exploring their structure, function and roles in disease pathogenesis and aging. We begin with an overview of mitochondrial complex I.
A diagrammatic representation of ETC complex I indicating its shape and function. Source: wikipedia.org
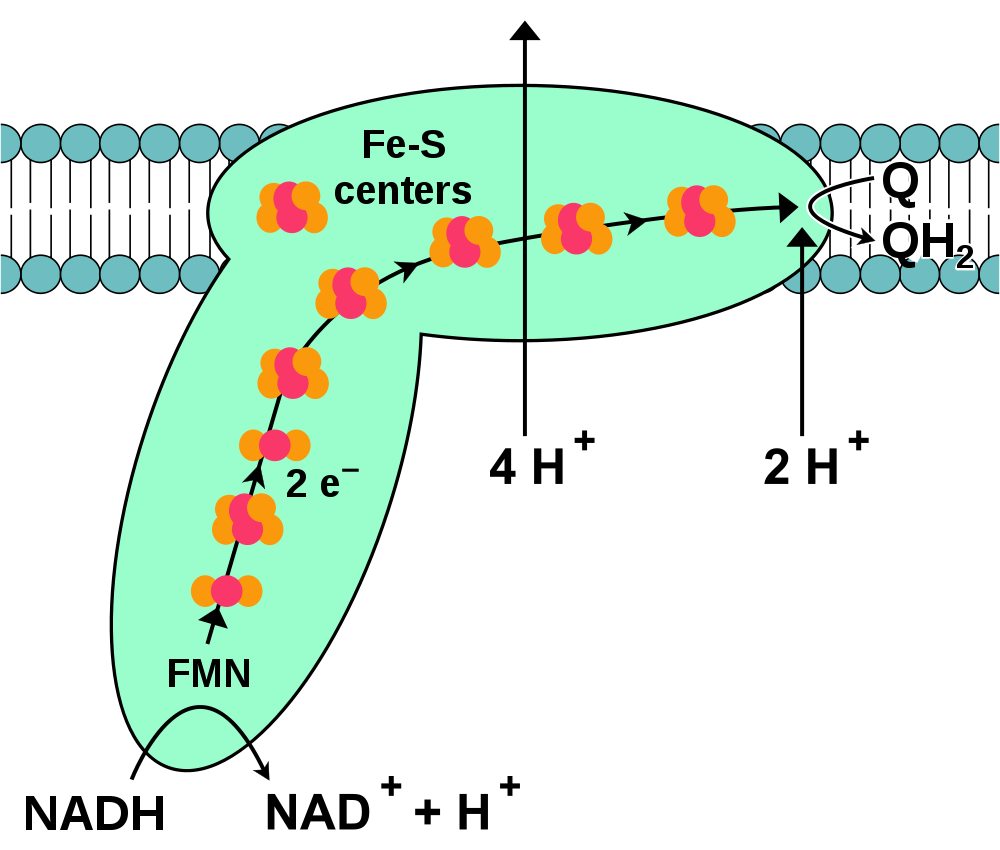
Complex I, or to give it its full name: NADH:ubiquinone oxidoreductase [Enzyme Commission number 1.6.5.3.] is an enzyme complex that catalyses the first step in the ETC. In mammals, complex I (CI) is composed of 44 sub-units [1]. The majority of these sub-units are encoded by the DNA contained within the nucleus of the cell (nuclear DNA or nDNA). However a small number of subunits are derived from the small circular DNA molecules contained within the mitochondria itself (mitochondrial DNA or mtDNA). Protein subunits encoded by both nDNA and mtDNA are needed to form the complete CI enzyme. Structurally, CI consists of two ‘arms’, one of which – the membrane arm- is embedded in the inner mitochondrial membrane, while the second – the hydrophilic arm – projects into the mitochondrial matrix [1].
The overall reaction catalysed by CI can be represented by the following chemical equation:
NADH + H+ + CoQ + 4H+in → NAD+ + CoQH2 + 4H+out
From the left hand side of this equation (and the diagram above) we can see that CI interacts with two substrates: nicotinamide adenine dinucleotide (NADH) and coenzyme Q10 (CoQ). This reaction also utilises a single free proton. Within the hydrophilic arm of CI is embedded a unit of flavin mononucleotide (FMN), Flavin mononucleotide (FMN), or riboflavin-5′-phosphate, is a molecule produced from riboflavin (vitamin B2) by the enzyme riboflavin kinase. FMN is a non-protein co-factor necessary for the biological activity of complex I. The reaction between NADH and FMN is the first step in the CI-mediated reaction [2] and serves to oxidise NADH in the following reaction, with a total of two electrons introduced into CI:
NADH + H+ + FMN → NAD+ + FMNH2
Following this initial oxidation, the two electrons liberated by FMN are passed to the first in a chain of iron-sulphur centres within the structure of CI [2]. This chain of molecules serves as a molecular ‘wire’ that connects FMN to a site in the CI membrane arm where the electrons are passed on to the inter-complex transport molecule CoQ (ubiquione) to form CoQH2 (ubiquinol) in the following step:
CoQ + 2H+ + 2e– → CoQH2
CoQ is highly hydrophobic, and as such it is found predominantly in the intermembrane space of the inner mitochondrial membrane. It is in this lipid-rich environment that CoQ is reduced to CoQH2 and through which it transports electrons to its target – complex III [3]. As well as its role in electron transport, the reduction of CoQ has an important second function; some of the energy released in this reaction is used to pump protons through CI from the mitochondrial matrix to the outside of the mitochondrial inner membrane thus generating an electrochemical proton gradient between the mitochondrial intermembrane space and the mitochondrial matrix. For each molecule of CoQH2 generated, 4 protons cross the inner membrane into the matrix [3]. It is this electrochemical gradient that underpins the ATP generating capability of ATP synthase.
Despite CI being essential for human life, it has also been implicated in several human disease states as well as in normal physiological aging. This is thought to be due to the fact that CI is a source of reactive oxygen species (ROS). ROS are molecules with ionic forms of oxygen that have been shown to contribute to cellular damage. ROS are generated as a by-product of the large amounts of oxygen that the mitochondrion consumes and there are mechanisms in place to neutralise these toxic molecules (such as superoxide dismutase, catalase and glutathione peroxidase as well as vitamin C , vitamin E, glutathione and uric acid). However ROS are sometimes produced at a rate greater than that which these mechanisms can operate [4], leading to a gradual build up of oxidative damage throughout life. The generation of ROS can be through a “vicious cycle”, as endogenously generated ROS can damage mitochondrial DNA (mtDNA), which in turn leads to aberrant production of CI (alongside other mitochondrial dysfunctions), which in turn leads to increased ROS generation [4]. As the various physiological effects of ROS-mediated mtDNA damage are compounded with age, they have been strongly implicated as a contributing factor to a range of diseases, namely so-called “mitochondrial diseases”, clinical presentations of which can include neurological, respiratory, gastrointestinal and endocrine conditions [5]. Despite their negative implications to health, it has been established that mitochondrial ROS production also plays an important role in normal cellular signalling and physiological regulation [6].
In short, CI liberates two electrons from NADH and transfers them to the transport molecule CoQ10 to be utilised further down the ETC. In this process an electrochemical proton gradient is generated by transporting 4 protons from the mitochondrial matrix into the mitochondrial intermembrane space. As a consequence of its function CI produces ROS that have been implicated in a range of disease states and in normal physiological aging. Our understanding of all of these mechanisms is incomplete, but there is an increasing feeling in the mitochondrial research community that we are on the threshold of being to positively manipulate and restore mitochondrial function/s in order to alleviate clinical conditions where mitochondrial dysfunction is a causative factor.
References and Further Reading:
1 – PDBe. Pumping protons: a complex problem. (link to article) (Accessed 7th May 2013)
2 – Vinogradov, A. D. (2008) NADH/NAD+ interaction of NADH:ubiquinone oxereductase (complex I). Biochimica et Biophysica Acta 1777(7-8) (link to article)
3 – Crane, F. L. (2001) Biochemical functions of coenzyme Q10. Journal of the American College of Nutrition 20(6) (link to article)
4 – Kirkinezos, I. G. & Moraes, C. T. (2001) Reactive oxygen species and mitochondrial diseases. Seminars in Cell & Developmental Biology 12 (link to article)
5 – Chial, H. & Craig, J. (2008) mtDNA and Mitochondrial diseases. Nature Education 1(1) (link to article)
6 – Sena, L. A. & Chandel, N. S. (2012) Physiological roles of mitochondrial reactive oxygen species. Molecular Cell 48(2) (link to article)

{kind=link}